In their environment, cells are subjected to various extracellular signals that orient their behavior by the activation of intracellular signaling cascades. In most cases, these signals are relayed by plasma membrane proteins whose activity and residence time at the membrane are finely regulated. Arrestins are part of these regulators. They constitute a superfamily of adaptor proteins present from protists to mammals, and share a common role in the regulation of the intracellular trafficking of membrane proteins and associated signaling.
The genome of the amoeba
Dictyostelium encodes six arrestin-like proteins (AdcA to F). In its natural habitat, this unicellular organism is able to respond to different chemoattractants such as folate released by the bacteria it feeds on and cAMP, secreted by the amoeba when faced with a lack of nutrients. Detection of these signals leads to the migration of cells towards the bacteria (folate) or to the aggregation of starving cells and the establishment of a multicellular developmental program leading to the differentiation of spores resistant to starvation (cAMP).
Researchers at the Large Scale Biology Laboratory and the Physiopathology of the Cytoskeleton Group have shown that the arrestin AdcC, known to regulate the cAMP receptor cAR1, is transiently recruited at the plasma membrane in response to cAMP and folate (Figure 1). They established that this response requires the calcium influx triggered by the chemoattractant-mediated activation of the receptors, and involves binding of calcium to the
C2 domaine of the AdcC protein and the oligomerization of AdcC
via its protein-protein interaction
SAM domains. Unexpectedly, AdcB which shares with AdcC the same structural organization and several biochemical properties including in vitro calcium binding, is insensitive to cAMP and folate, and does not respond to an increase in cytoplasmic calcium. These differences in behavior indicate different modes of regulation for these two close arrestins.
The data open interesting perspectives in the context of studies conducted on the pathogenic amoeba
Entamoeba histolytica responsible for amoebic dysentery in humans, that also harbors a C2 domain-containing arrestin. From an evolutionary point of view, such functional studies extended to other members of the arrestin family should allow the determination of ancestral functions supported by this family of proteins and the specificities added during evolution to meet the constraints related to the environment and the way of life of the different organisms.
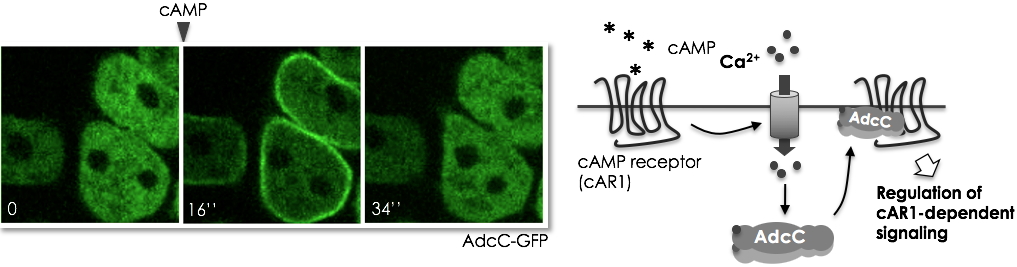
AdcC (in green) responds rapidly to a cAMP signal by a transient translocation of the cytosol to the plasma membrane (16’’ after stimulation) which reflects the intracellular calcium fluctuations triggered by the binding of the chemoattractant to its receptor.
Scheme of AdcC response to cAMP.
AdCC and AdcB arrestins, specific of Dictyostelids, have an arrestin core with a structure similar to that of mammalian β-arrestins, extended at the N-terminus by a calcium-dependent anionic lipid binding C2 domain, and at the C-terminal by a tandem SAM domain (sterile alpha domain).